O Transtorno do Espectro Autista (TEA) é amplamente reconhecido pela comunidade científica como uma das condições neurobiológicas com maior componente genético conhecido. Estudos epidemiológicos realizados com gêmeos e grandes coortes populacionais indicam que a herdabilidade do autismo — ou seja, a proporção da variação fenotípica que pode ser atribuída a fatores genéticos — situa-se entre 60% e 90%, podendo chegar a 98% em casos de gêmeos monozigóticos. Essa complexidade genética, no entanto, não segue as leis clássicas da herança mendeliana simples, como ocorre em doenças como a fibrose cística ou a anemia falciforme. Em vez disso, o TEA apresenta uma arquitetura genética multifatorial, onde a interação entre variantes genéticas comuns, mutações raras e fatores ambientais molda o desenvolvimento do sistema nervoso central.
A distinção entre o que é “genético” e o que é “hereditário” é fundamental para compreender quem “passa” o autismo. Enquanto o termo genético refere-se a qualquer alteração no DNA, o termo hereditário aplica-se especificamente às variações transmitidas de pais para filhos. No contexto do TEA, estima-se que a grande maioria dos casos tenha uma base genética, mas nem todos são herdados. Aproximadamente 80% do risco genético advém de variantes hereditárias, enquanto uma parcela significativa, especialmente em famílias sem histórico prévio da condição, resulta de mutações espontâneas que ocorrem nas células germinativas dos pais ou durante o desenvolvimento embrionário inicial.
Variantes Comuns e o Modelo Poligênico
A maior parte da arquitetura genética do autismo é composta por variantes genéticas comuns, conhecidas como Polimorfismos de Nucleotídeo Único (SNPs). Essas variantes estão presentes em uma parcela considerável da população geral e, individualmente, exercem um efeito muito pequeno sobre o fenótipo. No entanto, quando um indivíduo herda uma carga acumulada dessas variantes — o que os cientistas chamam de “escore de risco poligênico” —, o limiar de suscetibilidade para o autismo pode ser ultrapassado. Este modelo explica por que pais que não são autistas podem ter filhos no espectro: eles podem carregar subconjuntos dessas variantes que, ao se combinarem no descendente, resultam na manifestação do transtorno.
Essa herança poligênica é a razão pela qual traços de personalidade e comportamentos relacionados ao espectro, como foco intenso em detalhes ou dificuldades sutis de interação social, são frequentemente observados em parentes de primeiro grau de pessoas autistas, um fenômeno descrito como o “Fenótipo Ampliado do Autismo”. A combinação dessas variantes comuns é distribuída aleatoriamente durante a meiose, o que torna a manifestação do TEA um evento de probabilidade estatística dentro da dinâmica familiar.
Mutações Raras e Variantes de Novo
Diferente das variantes comuns, as variantes genéticas raras possuem um impacto funcional muito mais elevado. Elas podem incluir Variações no Número de Cópias (CNVs), que são deleções ou duplicações de grandes segmentos do DNA, ou mutações de ponto que alteram proteínas essenciais. Muitas dessas alterações são classificadas como mutações de novo, ou seja, não estão presentes no DNA somático dos pais, surgindo de forma espontânea.
As mutações de novo são particularmente prevalentes em casos de “famílias simplex”, onde há apenas um indivíduo autista em uma linhagem familiar sem histórico. Essas mutações frequentemente afetam genes envolvidos em processos críticos, como a formação de sinapses e a regulação da expressão gênica no cérebro. A ocorrência dessas mutações espontâneas desafia a noção de que o autismo é sempre “passado” linearmente, demonstrando que a biologia reprodutiva humana possui uma margem intrínseca de variabilidade que pode resultar em neurodivergência independentemente da herança parental direta.
O Papel da Idade Paterna e a Contribuição dos Genitores
A questão de quem “passa” o autismo frequentemente recai sobre a análise das contribuições paterna e materna. Estudos genômicos indicam que a idade avançada do pai é um dos fatores de risco ambientais-genéticos mais consistentes para a ocorrência de mutações de novo. À medida que os homens envelhecem, as células precursoras dos espermatozoides passam por sucessivas divisões celulares, aumentando a probabilidade de erros na replicação do DNA. Isso resulta em um maior número de mutações pontuais transmitidas pela linhagem germinativa masculina.
Por outro lado, a contribuição materna é igualmente complexa. Embora as mulheres pareçam ter um “efeito protetor” biológico contra a manifestação do autismo — necessitando de uma carga genética muito maior para apresentar o fenótipo do que os homens —, elas podem ser portadoras assintomáticas de variantes raras de alto impacto. Nesses casos, a mãe pode transmitir uma variante para o filho que, devido à ausência do efeito protetor feminino no sexo masculino, resulta em um diagnóstico de TEA. Portanto, a transmissão hereditária pode ocorrer de ambos os progenitores, mas os mecanismos de expressão fenotípica variam significativamente entre os sexos.
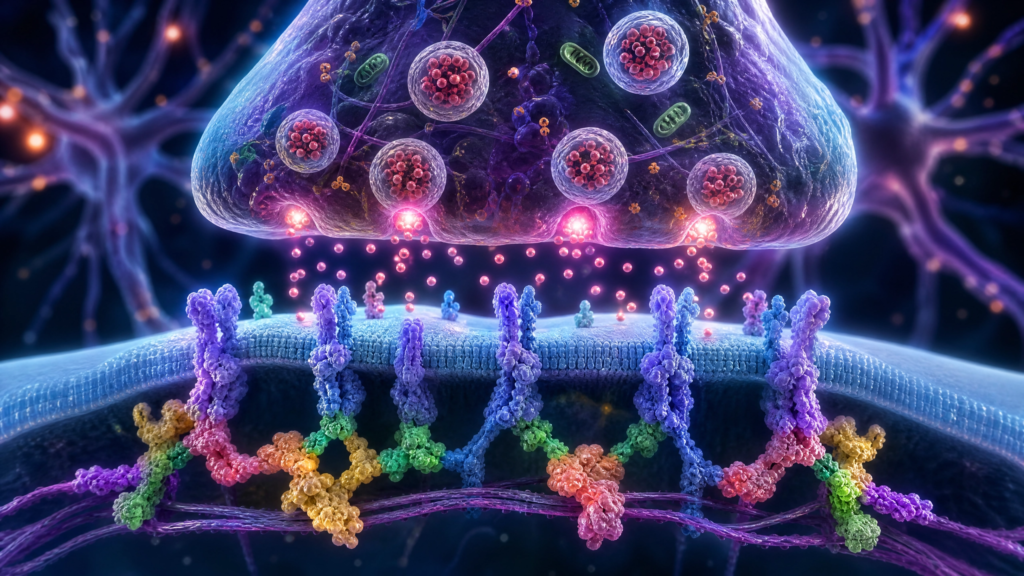
Mecanismos Biológicos: Sinapses e Conectividade Neural
A nível molecular, os genes associados ao TEA convergem para vias biológicas específicas que ditam como os neurônios se comunicam. Um dos grupos de genes mais estudados é o das proteínas de adesão celular sináptica, como as Neuroliginas (NLGN3, NLGN4X) e as Neurexinas (NRXN1). Essas proteínas funcionam como “pontes” que conectam os neurônios nas sinapses, garantindo que os sinais químicos sejam transmitidos de forma eficiente. Alterações nesses genes perturbam o equilíbrio entre a excitação e a inibição neuronal, uma característica central da neurofisiologia do autismo.
Outro gene de destaque é o SHANK3, localizado no cromossomo 22. Ele codifica uma proteína de andaime na densidade pós-sináptica que organiza centenas de outras proteínas necessárias para a plasticidade sináptica. Mutações ou deleções no SHANK3 estão fortemente ligadas ao autismo e a síndromes relacionadas, como a Síndrome de Phelan-McDermid. Além da função sináptica, muitos genes do TEA estão envolvidos na remodelação da cromatina e na regulação da transcrição, o que significa que eles controlam quando e como outros genes são ativados durante o desenvolvimento embrionário do cérebro.
Epigenética e a Interação com o Ambiente
A genética do autismo não é um destino selado apenas pela sequência de letras do DNA (A, T, C, G). A epigenética — o estudo de mudanças químicas que ativam ou desativam genes sem alterar a sequência do DNA — desempenha um papel crucial. Fatores como a metilação do DNA e a modificação de histonas podem ser influenciados pelo ambiente intrauterino, incluindo o estado imunológico da mãe, a exposição a certas substâncias químicas e níveis hormonais.
Essas marcas epigenéticas podem determinar se um gene de risco herdado será expresso ou silenciado. Isso ajuda a explicar a discordância fenotípica em gêmeos idênticos, onde um pode ser autista e o outro não, ou apresentar níveis de suporte diferentes. A interação gene-ambiente sugere que o autismo é o resultado de uma predisposição biológica que responde de maneira única aos estímulos e condições externas durante períodos críticos do desenvolvimento neuroembriológico.
O Efeito Protetor Feminino e o Viés de Gênero
A prevalência do autismo é notavelmente maior em homens, com uma proporção de aproximadamente 4 para 1. A genética oferece uma explicação robusta para essa disparidade através da teoria do “Efeito Protetor Feminino”. Pesquisas mostram que mulheres autistas tendem a carregar um número significativamente maior de mutações deletérias e variações no número de cópias do que homens autistas com o mesmo nível de gravidade de sintomas.
Isso sugere que o cérebro feminino possui mecanismos de resiliência — possivelmente ligados a fatores hormonais como o estrógeno ou à presença de dois cromossomos X — que exigem um “golpe” genético muito mais forte para que o TEA se manifeste clinicamente. Como consequência, homens com uma carga genética moderada podem ser diagnosticados, enquanto mulheres com a mesma carga podem permanecer abaixo do limiar clínico, embora possam transmitir essa carga genética para seus descendentes.
Síndromes Monogênicas e Diagnóstico Diferencial
Embora a maioria dos casos de TEA seja idiopática (de causa multifatorial desconhecida), cerca de 10% a 15% dos diagnósticos estão associados a síndromes genéticas bem definidas. A Síndrome do X Frágil, causada por uma mutação no gene FMR1, é a causa hereditária mais comum de deficiência intelectual e autismo em homens. Outras condições incluem a Síndrome de Rett (gene MECP2), a Esclerose Tuberosa (genes TSC1 e TSC2) e a Síndrome de Angelman.
Identificar essas causas específicas através de exames como o Microarray (CMA) ou o Sequenciamento Completo do Exoma (WES) é vital para o manejo clínico. Essas condições frequentemente trazem comorbidades médicas específicas, como epilepsia, problemas cardíacos ou distúrbios metabólicos, que requerem monitoramento direcionado. O diagnóstico molecular, portanto, transforma a compreensão do autismo de uma observação puramente comportamental para uma análise biológica precisa, permitindo que as famílias compreendam a origem exata da condição e os riscos de recorrência em futuras gestações.
A Complexidade do Aconselhamento Genético
Dada a natureza heterogênea do TEA, o aconselhamento genético tornou-se uma ferramenta indispensável. Ele não busca apenas identificar “quem passou”, mas sim oferecer uma visão clara da arquitetura biológica do indivíduo. Quando uma mutação de novo é identificada, o risco de recorrência para o próximo filho do casal é geralmente baixo (cerca de 1% a 2%, devido ao mosaicismo germinativo). No entanto, se o autismo na família for predominantemente impulsionado por uma alta carga de variantes comuns ou por uma variante rara herdada, o risco de recorrência pode ser significativamente maior, chegando a 20% ou mais em algumas linhagens.
O avanço das tecnologias de sequenciamento permitiu a identificação de mais de 100 genes de alto risco e centenas de outros candidatos. Essa lista continua a crescer à medida que estudos globais integram dados de milhares de genomas. A ciência moderna caminha para uma compreensão onde o autismo não é visto como um erro, mas como uma manifestação da vasta diversidade genética humana. A “passagem” do autismo, portanto, é um processo intrincado de herança de predisposições, ocorrências aleatórias de mutações e influências ambientais que, juntos, resultam na singularidade do desenvolvimento neurodivergente.
| Categoria de Variante | Origem | Impacto Fenotípico | Frequência na População |
| Variantes Comuns (SNPs) | Hereditária (Ambos os pais) | Pequeno (Efeito cumulativo) | Alta |
| Mutações De Novo | Espontânea (Geralmente linhagem paterna) | Alto | Rara |
| CNVs (Deleções/Duplicações) | Hereditária ou De Novo | Moderado a Alto | Rara |
| Síndromes Monogênicas | Hereditária (Ex: X-Frágil) | Muito Alto | Muito Rara |
A análise da genética do TEA revela que a pergunta “quem passa o autismo” não possui uma resposta única. Em alguns casos, é uma herança direta de variantes raras; em outros, é o resultado de uma loteria genética de variantes comuns que se alinharam de forma específica; e em muitos outros, é um evento biológico novo e espontâneo. O que permanece constante é a evidência de que o autismo está profundamente enraizado na biologia humana, refletindo a complexidade da evolução do nosso cérebro e a interação dinâmica entre o código genético e o mundo em que vivemos.